
Recommendation
Suidae are well-represented in Plio-Pleistocene African hominin sites and are particularly important for biochronological assessments. Their ubiquity in hominin sites combined with multiple appearances of what appears to be graminivorous adaptations in the lineage (Harris & White, 1979) suggest that they have the potential to contribute to our understanding of Plio-Pleistocene paleoenvironments. While they have been generally understudied in this respect, there has been recent focus on their diets to understand the paleoenvironments of early hominin habitats. Of particular interest is Kolpochoerus, one of the most abundant suid genera in the Plio-Pleistocene with a wide geographic distribution and diverse dental morphologies (Harris & White, 1979).
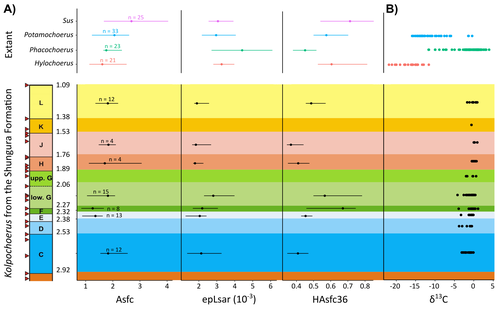
In this study, Louail et al. (2024) present the results of the first dental microwear texture analysis (DMTA) conducted on suids from the Shungura Formation of the Omo Valley, an important Plio-Pleistocene hominin site that records an almost continuous sedimentary record from ca. 3.75 Ma to 1.0 Ma (Heinzelin 1983; McDougall et al., 2012; Kidane et al., 2014). Dental microwear is one of the main proxies in understanding diet in fossil mammals, particularly herbivores, and DMTA has been shown to be effective in differentiating inter- and intra-species dietary differences (e.g., Scott et al., 2006; 2012; Merceron et al., 2010). However, only a few studies have applied this method to extinct suids (Souron et al., 2015; Ungar et al., 2020), making this study especially pertinent for those interested in suid dietary evolution or hominin paleoecology.
In addition to examining DMT variations of Kolpochoerus specimens from Omo, Louail et al. (2024) also expanded the modern comparative data set to include larger samples of African suids with different diets from herbivores to omnivores to better interpret the fossil data. They found that DMTA distinguishes between extant suid taxa, reflecting differences in diet, indicating that DMT can be used to examine the diets of fossil suids. The results suggest that Kolpochoerus at Omo had a substantially different diet from any extant suid taxon and that although its anistropy values increased through time, they remain well below those observed in modern Phacochoerus that specializes in fibrous, abrasive plants. Based on these results, in combination with comparative and experimental DMT, enamel carbon isotopic, and morphological data, Louail et al. (2024) propose that Omo Kolpochoerus preferred short, soft and low abrasive herbaceous plants (e.g., fresh grass shoots), probably in more mesic habitats. Louail et al. (2024) note that with the wide temporal and geographic distribution of Kolpochoerus, different species and populations may have had different feeding habits as they exploited different local habitats. However, it is noteworthy that similar inferences were made at other hominin sites based on other types of dietary data (e.g., Harris & Cerling, 2002; Rannikko et al., 2017, 2020; Yang et al., 2022). If this is an indication of their habitat preferences, the wide-ranging distribution of Kolpochoerus may suggest that mesic habitats with short, soft herbaceous plants were present in various proportions at most Plio-Pleistocene hominin sites.
References
Harris, J. M., and Cerling, T. E. (2002). Dietary adaptations of extant and Neogene African suids. Journal of Zoology, 256(1), 45–54. https://doi.org/10.1017/S0952836902000067
Harris, J. M., and White, T. D. (1979). Evolution of the Plio-Pleistocene African Suidae. Transactions of the American Philosophical Society, 69(2), 1–128. https://doi.org/10.2307/1006288
Heinzelin, J. de. (1983). The Omo Group. Archives of the International Omo Research Expedition. Volume 85. Annales du Musée Royal de l’Afrique Centrale, série 8, Sciences géologiques, Tervuren, 388 p.
Kidane, T., Brown, F. H., and Kidney, C. (2014). Magnetostratigraphy of the fossil-rich Shungura Formation, southwest Ethiopia. Journal of African Earth Sciences, 97, 207–223. https://doi.org/10.1016/j.jafrearsci.2014.05.005
Louail, M., Souron, A., Merceron, G., and Boisserie, J.-R. (2024). New insights on feeding habits of Kolpochoerus from the Shungura Formation (Lower Omo Valley, Ethiopia) using dental microwear texture analysis. PaleorXiv, dbgtp, ver. 3, peer-reviewed by PCI Paleo. https://doi.org/10.31233/osf.io/dbgtp
McDougall, I., Brown, F. H., Vasconcelos, P. M., Cohen, B. E., Thiede, D. S., and Buchanan, M. J. (2012). New single crystal 40Ar/39Ar ages improve time scale for deposition of the Omo Group, Omo–Turkana Basin, East Africa. Journal of the Geological Society, 169(2), 213–226. https://doi.org/10.1144/0016-76492010-188
Merceron, G., Escarguel, G., Angibault, J.-M., and Verheyden-Tixier, H. (2010). Can dental microwear textures record inter-individual dietary variations? PLoS ONE, 5(3), e9542. https://doi.org/10.1371/journal.pone.0009542
Rannikko, J., Adhikari, H., Karme, A., Žliobaitė, I., and Fortelius, M. (2020). The case of the grass‐eating suids in the Plio‐Pleistocene Turkana Basin: 3D dental topography in relation to diet in extant and fossil pigs. Journal of Morphology, 281(3), 348–364. https://doi.org/10.1002/jmor.21103
Rannikko, J., Žliobaitė, I., and Fortelius, M. (2017). Relative abundances and palaeoecology of four suid genera in the Turkana Basin, Kenya, during the late Miocene to Pleistocene. Palaeogeography, Palaeoclimatology, Palaeoecology, 487, 187–193. https://doi.org/10.1016/j.palaeo.2017.08.033
Scott, R. S., Teaford, M. F., and Ungar, P. S. (2012). Dental microwear texture and anthropoid diets. American Journal of Physical Anthropology, 147(4), 551–579. https://doi.org/10.1002/ajpa.22007
Scott, R. S., Ungar, P. S., Bergstrom, T. S., Brown, C. A., Childs, B. E., Teaford, M. F., and Walker, A. (2006). Dental microwear texture analysis: Technical considerations. Journal of Human Evolution, 51(4), 339–349. https://doi.org/10.1016/j.jhevol.2006.04.006
Souron, A., Merceron, G., Blondel, C., Brunetière, N., Colyn, M., Hofman-Kamińska, E., and Boisserie, J.-R. (2015). Three-dimensional dental microwear texture analysis and diet in extant Suidae (Mammalia: Cetartiodactyla). Mammalia, 79(3). https://doi.org/10.1515/mammalia-2014-0023
Ungar, P. S., Abella, E. F., Burgman, J. H. E., Lazagabaster, I. A., Scott, J. R., Delezene, L. K., Manthi, F. K., Plavcan, J. M., and Ward, C. V. (2020). Dental microwear and Pliocene paleocommunity ecology of bovids, primates, rodents, and suids at Kanapoi. Journal of Human Evolution, 140, 102315. https://doi.org/10.1016/j.jhevol.2017.03.005
Yang, D., Pisano, A., Kolasa, J., Jashashvili, T., Kibii, J., Gomez Cano, A. R., Viriot, L., Grine, F. E., and Souron, A. (2022). Why the long teeth? Morphometric analysis suggests different selective pressures on functional occlusal traits in Plio-Pleistocene African suids. Paleobiology, 48(4), 655–676. https://doi.org/10.1017/pab.2022.11
DOI or URL of the preprint: https://doi.org/10.31233/osf.io/dbgtp
Version of the preprint: 1
Dear Denise Su,
We thank you and the two reviewers for revising the initial version of our manuscript.
We have made every attempt to fully address the constructive comments outlined by you and the reviewers. We believe these revisions have strongly improved the initial version of our manuscript. Our replies can be found directly in the provided pdf.
Particularly, we have provided clarifications on the materials and methods, in the main text as in the supplementary material (ESM1: Tables S1 to S3). Tooth positions are specified (ESM1: Tables S1 and S2). We also provide additional tests showing that DMT do not significantly differ between tooth position, as well as references to previous studies showing no significant difference between tooth position when considering homologous wear facets. Overall, this justifies the inclusion of all suitable molars (worn, with non-altered facets) in the analyses.
We have followed suggestions made by the reviewers to improve figure and text clarity. We have combined Figures 3 and 4 into Figure 3, as well as Figures 4 and 5 into Figure 4. We think that including in the same figure both the data at the generic and specific levels (median and interquartile ranges), as well as individual data points (including data by members for Kolpochoerus) resulted in an overloaded and not easily readable figure. However, it is indeed important to show individual data points. Thus, we have included an additional figure in the supplementary in that sense (ESM1: Figure S1).
At last, we have included a discussion point to contextualize the results of the present study in relation to the wide distribution of Kolpochoerus in Plio-Pleistocene Africa and its dietary versatility.
We hope that the revised version of our manuscript will meet the reviewers’ expectations as well as yours.
Sincerely yours,
Margot Louail
This is an excellent, well thought-out study that would add valuable data for interpreting early hominin habitats as suids are common and certainly Kolpochoerus is one of the most wide ranging taxon. Based on reviewer comments and my own reading of the manuscript, I believe that it would benefit from clarifications on the materials and methods. Both reviewers and I noted that it appears that different teeth are lumped in the analyses and wonder about its impact on the results and interpretation. The reviewers also provide suggestions on improving figure and text clarity, which I would recommend that the authors consider. I am, personally, particularly interested in the question of the dietary versatility of Kolpochoerus in its wide distribution. Of course, there were likely differences for each species, but it would be an interesting discussion point given how ubiquitous it is in Plio-Pleistocene Africa. I understand that this is a manuscript focused on the Omo, but a discussion to contextualize the results, particularly in terms of dietary trends, would be helpful.
 , 20 Dec 2023
, 20 Dec 2023Louail and colleagues provide an interesting contribution to dietary ecology and dental wear in both extant suids and the extinct genus Kolpochoerus by using dental microwear texture analysis – a wearbased proxy that evaluates microscopic wear patterns on enamel using semi-automated, objective quantification of these wear marks. Their data expands available dataset of extant suids, thus both contributing to our understanding of how extant dietary diversity is reflected in dental wear, and on how Kolpochoerus from the Omo Valley differed in its diet from extant counterparts. This kind of study is also interesting, because it allows a better understanding of the palaeohabitat at the Lower Omo valley, as habitat and food availability are directly connected. It is, of course, still a local snapshot of only one species from Lower Omo valley, and specific to suids, but still the potential to derive general habitat reconstruction is given.
Therefore, I would assess the novelty and impact of this study positively. There are a few points that need attention, especially design of the figures and selection of reference, and the composition of the dataset. It is unclear which tooth positions were exactly used, and if lumping together different tooth position is justified and does not weaken the analysis. But I am positive that the authors will be able to account for them. If I were to give a suggestion for acceptance in a journal, it would be “minor revisions”.
See specific remarks in the attached PDF.
Download the review https://doi.org/10.24072/pci.paleo.100255.rev11I have attached the manuscript with track changes and comments. The majority of my suggested changes were simply grammatical/translational issues. Overall, this was an excellent and well-thought out paper. I was particularly pleased to see Figure 2's visual clarifications of the types of surface damage that was omitted and found the ecological considerations i the discussion to be comprehensive. My one concern is that data was obtained from the M1-3 and m1-3, no doubt to increase the sample sizes across members, but in doing so without listing them in the supplement makes it unreplicable and also possibly muddles the data. Was it only specimens included were all six of these molars were available? The M3 is typically used, so how does the inclusion of data from other less specialzied molars impact the resulting data? I ask that this is better adressed, including the addition of which molars were used for each of the included specimens in the supplement.
Download the review https://doi.org/10.24072/pci.paleo.100255.rev12