
Latest recommendations
| Id | Title * | Authors * | Abstract * | Picture * | Thematic fields * | Recommender | Reviewers | Submission date | |
|---|---|---|---|---|---|---|---|---|---|
27 May 2020

The last surviving Thalassochelydia—A new turtle cranium from the Early Cretaceous of the Purbeck Group (Dorset, UK)A recommendation of: The last surviving Thalassochelydia—A new turtle cranium from the Early Cretaceous of the Purbeck Group (Dorset, UK)Recommended by Hans-Dieter Sues based on reviews by Igor Danilov and Serjoscha EversStem- and crown-group turtles have a rich and varied fossil record dating back to the Triassic Period. By far the most common remains of these peculiar reptiles are their bony shells and fragments of shells. Furthermore, if historical specimens preserved skulls the preparation techniques at that time were inadequate for elucidating details of the cranial structure. Thus, it comes as no surprise that most of the early research on turtles focused on the structure of the shell with little attention paid to other parts of the skeleton. Starting in the 1960s, this changed as researchers realized that there is considerable variation in the structure of turtle shells even within species and that new methods of fossil preparation, especially chemical methods, could reveal a wealth of phylogenetically important features in the structure of the skulls of turtles. The principal worker was Eugene S. Gaffney of the American Museum of Natural History (New York) who in a series of exquisitely illustrated monographs revolutionized our understanding of turtle osteology and phylogeny. Over the last decade or so, a new generation of researchers has further refined the phylogenetic framework for turtles and continued the work by Gaffney. One of the specialists from this new generation is Jérémy Anquetin who, with a number of colleagues, has revised many of the Jurassic-age stem-turtles that existed in coastal marine settings in what is now Europe. Collections in France, Germany, Switzerland, and the UK house numerous specimens of these forms, which attracted the interest of researchers as early as the first decades of the nineteenth century. Despite this long history, however, the diversity and interrelationships of these marine taxa remained poorly understood. In the present study, Anquetin and his colleague Charlotte André extend the fossil record of these stem-turtles, recently hypothesized as a clade Thalassochelydia, into the Early Cretaceous (Anquetin & André 2020). They present an excellent anatomical account on a well-preserved cranium from the Purbeck Formation of Dorset (England) that can be referred to Thalassochelydia and augments our knowledge of the cranial morphology of this clade. Anquetin & André (2020) make a good case that this specimen belongs to the same taxon as shell material long ago described as Hylaeochelys belli. References Anquetin, J., & André, C. (2020). The last surviving Thalassochelydia—A new turtle cranium from the Early Cretaceous of the Purbeck Group (Dorset, UK). PaleorXiv, 7pa5c, version 3, peer-reviewed by PCI Paleo. doi: 10.31233/osf.io/7pa5c | The last surviving Thalassochelydia—A new turtle cranium from the Early Cretaceous of the Purbeck Group (Dorset, UK) | Jérémy Anquetin, Charlotte André | <p>**Background.** The mostly Berriasian (Early Cretaceous) Purbeck Group of southern England has produced a rich turtle fauna dominated by the freshwater paracryptodires *Pleurosternon bullockii* and *Dorsetochelys typocardium*. Each of these spe... | | Comparative anatomy, Paleoecology, Phylogenetics, Systematics, Vertebrate paleontology | Hans-Dieter Sues | 2020-01-30 10:37:07 | ||
27 Jan 2020

A simple generative model of trilobite segmentation and growthDeep insights into trilobite developmentRecommended by Christian Klug based on reviews by Kenneth De Baets and Lukas LaiblTrilobites are arthropods that became extinct at the greatest marine mass extinction over 250 Ma ago. Because of their often bizarre forms, their great diversity and disparity of shapes, they have attracted the interest of researchers and laypersons alike. Due to their calcified exoskeleton, their remains are quite abundant in many marine strata. One particularly interesting aspect, however, is the fossilization of various molting stages. This allows the reconstruction of both juvenile strategies (lecitotrophic versus planktotrophic) and the entire life history of at least some well-documented taxa (e.g., Hughes 2003, 2007; Laibl 2017). For example, life history of trilobites is, based on certain morphological changes, classically subdivided in the three phases protaspis (hatchling, one dorsal shield with few segments with no articulation between), meraspis (juvenile, two and more shields connected by articulations) and holaspis (when the terminal number of thoracic segments is reached). At most molting events, a new skeletal element is added (only in the holaspis, the number of thoracic segments does not change). Nevertheless, many trilobites are known mainly from late meraspid and holaspid stages, because the dorsal shields of the first ontogenetic stages are usually very small and thus often either dissolved or overlooked. An improved understanding of trilobite ontogeny could thus help filling in these gaps in fossil preservation and subsequently, to better understand evolutionary pathways. This is where this paper comes in. In a very clever approach, the New-York-based researcher Melanie Hopkins modeled the growth of these segmented animals (Hopkins 2020). Previous growth models of invertebrates focused on, e.g., mollusks, whose shells grow by accretion. Modelling arthropod ontogeny represented a challenge, which is now overcome by Hopkins' brilliant paper. Her generative growth model is based on empirical data of *Aulacopleura koninckii* (Barrande, 1846). Hong et al. (2014) and Hughes et al. (2017) documented the ontogeny of this 429 Ma old trilobite species in great detail. In the Silurian of the Barrandian region (Czech Republic), this species is locally very common and all growth stages are well known. I could imagine that the paper of Hughes et al. (2017) planted the seed into Melanie Hopkins’ mind to approach trilobite development in general in a quantitative way with a mathematical approach comparable to the mollusk-research by, e.g., David Raup (1961, 1966) and George McGhee (2015). Hopkins’ growth model requires “a minimum of nine parameters […] to model basic trilobite growth and segmentation, and three additional parameters […] to allow a transition to a new growth gradient for the trunk region during ontogeny” (Hopkins 2020: p. 21). It is now possible to play with parameters such as protaspid size, segment dimensions, segment numbers, etc., in order to estimate changes in body size or morphology. Furthermore, the model could be applied to similarly organized arthropod exoskeletons like many early Cambrian arthropods (e.g., marellomorphs) or even crustaceans (e.g., conchostracans or copepods). Of great interest could also be to assess influences of environmental changes on arthropod ontogeny. Also, her work will help to reconstruct unknown developmental information missing from trilobite species (and possibly other arthropods) and also to explore their morphospace. **References** Barrande, J. (1846). Notice préliminaire sur le système Silurien et les trilobites de Bohême. Leipzig: Hirschfield. Hong, P. S., Hughes, N. C., & Sheets, H. D. (2014). Size, shape, and systematics of the Silurian trilobite *Aulacopleura koninckii*. Journal of Paleontology, 88(6), 1120–1138. doi: [ 10.1666/13-142](https://dx.doi.org/ 10.1666/13-142) Hopkins, M. J. (2020). A simple generative model of trilobite segmentation and growth. PaleorXiv, version 3, peer-reviewed by PCI Paleo. doi: [ 10.31233/osf.io/zt642](https://dx.doi.org/ 10.31233/osf.io/zt642) Hughes, N. C. (2003). Trilobite tagmosis and body patterning from morphological and developmental perspectives. Integrative and Comparative Biology, 43(1), 185–206. doi: [ 10.1093/icb/43.1.185](https://dx.doi.org/ 10.1093/icb/43.1.185) Hughes, N. C. (2007). The evolution of trilobite body patterning. Annual Review of Earth and Planetary Sciences, 35(1), 401–434. doi: [ 10.1146/annurev.earth.35.031306.140258](https://dx.doi.org/ 10.1146/annurev.earth.35.031306.140258) Hughes, N. C., Hong, P. S., Hou, J., & Fusco, G. (2017). The development of the Silurian trilobite *Aulacopleura koninckii* reconstructed by applying inferred growth and segmentation dynamics: A case study in paleo-evo-devo. Frontiers in Ecology and Evolution, 5, 00037. doi: [ 10.3389/fevo.2017.00037](https://dx.doi.org/ 10.3389/fevo.2017.00037) Laibl, L. (2017). Patterns in Palaeontology: The development of trilobites. Palaeontology Online, 7(10), 1–9. McGhee, G. R. (2015). Limits in the evolution of biological form: a theoretical morphologic perspective. Interface Focus, 5(6), 20150034. doi: [ 10.1098/rsfs.2015.0034](https://dx.doi.org/ 10.1098/rsfs.2015.0034) Raup, D. M. (1961). The geometry of coiling in gastropods. Proceedings of the National Academy of Sciences, 47(4), 602–609. doi: [ 10.1073/pnas.47.4.602](https://dx.doi.org/ 10.1073/pnas.47.4.602) Raup, D. M. (1966). Geometric analysis of shell coiling: general problems. Journal of Paleontology, 40, 1178–1190. | A simple generative model of trilobite segmentation and growth | Melanie J Hopkins | <p>Generative growth models have been the basis for numerous studies of morphological diversity and evolution. Most work has focused on modeling accretionary growth systems, with much less attention to discrete growth systems. Generative growth mo... | | Evo-Devo, Evolutionary biology, Invertebrate paleontology, Paleobiology | Christian Klug | 2019-10-06 00:27:25 | ||
30 Oct 2019

The Morrison Formation Sauropod Consensus: A freely accessible online spreadsheet of collected sauropod specimens, their housing institutions, contents, references, localities, and other potentially useful informationSauropods under one (very high) roofRecommended by Jordan Mallon based on reviews by Kenneth Carpenter and Femke HolwerdaFossils get around. Any one fossil locality might be sampled by several collectors from as many institutions around the world. Alternatively, a single collector might heavily sample a site, and sell or trade parts of their collection to other institutions, scattering the fossils far and wide. These practices have the advantage of making fossils from any one locality available to researchers across the globe. However, they also have the disadvantage that, in order to systematically survey any one species, a researcher must follow innumerable trails of breadcrumb to get to where the relevant materials are held. This is true of many famous fossil localities, such as the Eocene Green River Formation in the USA, the Cretaceous Kem Kem beds of Morocco, or the Devonian Miguasha cliffs of Canada. It is especially true of the Upper Jurassic deposits of the Morrison Formation in the western USA, which have yielded an impressive assemblage of megaherbivorous sauropod dinosaurs over the last 150 years. Today, these bones are to be found in museums not just in the USA, but also in Canada, Argentina, Japan, Australia, Malaysia, South Africa, and throughout Europe. Trawling museum databases in search of sauropod material from the Morrison Formation can therefore be a daunting task, never mind traveling the globe to actually study them. A new paper by Tschopp et al. (2019) seeks to ease the burden on sauropod researchers by introducing a database of Morrison Formation sauropods, consisting of over 3000 specimens housed in nearly 40 institutions around the world. The authors are themselves sauropod workers and, having suffered first-hand the plight of studying material from the Morrison Formation, came up with a solution to the problem of keeping track of it all. The database is founded largely on material personally seen by the authors, supplemented by information from the literature and museum catalogs. The database further provides information on bone representation, ontogeny, locality details, and fine-scale stratigraphy, among other fields. Like any database, it is a living document that will continue to grow as new finds are made. Tschopp et al. (2019) have wisely chosen to allow others to contribute to the listing, but changes must first be vetted for accuracy. This product represents 10 years of work, and I have little doubt that it will be well-received by those of us who work on dinosaurs. Speaking personally, my PhD research on megaherbivorous dinosaurs from the Dinosaur Park Formation of Canada led me to institutions in Canada, the USA, and the UK, and further stops to Spain and Argentina would have been beneficial, if affordable. Planning for this work would have been greatly assisted by a database like the one provided us by Tschopp et al. (2019). Many a future graduate student will undoubtedly owe them a debt of gratitude. References Tschopp, E., Whitlock, J. A., Woodruff, D. C., Foster, J. R., Lei, R., & Giovanardi, S. (2019). The Morrison Formation Sauropod Consensus: A freely accessible online spreadsheet of collected sauropod specimens, their housing institutions, contents, references, localities, and other potentially useful information. PaleorXiv, version 3, peer-reviewed by PCI Paleo. doi: 10.31233/osf.io/ydvra | The Morrison Formation Sauropod Consensus: A freely accessible online spreadsheet of collected sauropod specimens, their housing institutions, contents, references, localities, and other potentially useful information | Emanuel Tschopp, John A. Whitlock, D. Cary Woodruff, John R. Foster, Roberto Lei, Simone Giovanardi | <p>The Morrison Formation has been explored for dinosaurs for more than 150 years, in particular for large sauropod skeletons to be mounted in museum exhibits around the world. Several long-term campaigns to the Jurassic West of the United States ... | | Fossil record, Methods, Paleobiodiversity, Taxonomy, Vertebrate paleontology | Jordan Mallon | 2019-07-19 16:13:45 | ||
16 Oct 2019

What do ossification sequences tell us about the origin of extant amphibians?The origins of LissamphibiaRecommended by Robert Asher ? based on reviews by Jennifer Olori and 2 anonymous reviewersAmong living vertebrates, there is broad consensus that living tetrapods consist of amphibians and amniotes. Crown clade Lissamphibia contains frogs (Anura), salamanders (Urodela) and caecilians (Gymnophiona); Amniota contains Sauropsida (reptiles including birds) and Synapsida (mammals). Within Lissamphibia, most studies place frogs and salamanders in a clade together to the exclusion of caecilians (see Pyron & Wiens 2011). Among fossils, there are a number of amphibian and amphibian-like taxa generally placed in Temnospondyli and Lepospondyli. In contrast to the tree of living tetrapods, affinities of these fossils to some or all of the three extant lissamphibian groups have proven to be much harder to resolve. For example, temnospondyls might be stem tetrapods and lissamphibians a derived group of lepospondyls; alternatively, temnospondyls might be closer to the clade of frogs and salamanders, and lepospondyls to caecilians (compare Laurin et al. 2019: fig. 1d vs. 1f). Here, in order to assess which of these and other mutually exclusive topologies is optimal, Laurin et al. (2019) extract phylogenetic information from developmental sequences, in particular ossification. Several major differences in ossification are known to distinguish vertebrate clades. For example, due to their short intrauterine development and need to climb from the reproductive tract into the pouch, marsupial mammals famously accelerate ossification of their facial skeleton and forelimb; in contrast to placentals, newborn marsupials can climb, smell & suck before they have much in the way of lungs, kidneys, or hindlimbs (Smith 2001). Divergences among living and fossil amphibian groups are likely pre-Triassic (San Mauro 2010; Pyron 2011), much older than a Jurassic split between marsupials and placentals (Tarver et al. 2016), and the quality of the fossil record generally decreases with ever-older divergences. Nonetheless, there are a number of well-preserved examples of "amphibian"-grade tetrapods representing distinct ontogenetic stages (Schoch 2003, 2004; Schoch and Witzmann 2009; Olori 2013; Werneburg 2018; among others), all amenable to analysis of ossification sequences. Putting together a phylogenetic dataset based on ossification sequences is not trivial; sequences are not static features apparent on individual specimens. Rather, one needs multiple specimens representing discrete developmental stages for each taxon to be compared, meaning that sequences are usually available for only a few characters. Laurin et al. (2019) have nonetheless put together the most exhaustive matrix of tetrapod sequences so far, with taxon coverage ranging from 62 genera for appendicular characters to 107 for one of their cranial datasets, each sampling between 4-8 characters (Laurin et al. 2019: table 1). The small number of characters means that simply applying an optimality criterion (such as parsimony) is unlikely to resolve most nodes; treespace is too flat to be able to offer optimal peaks up which a search algorithm might climb. However, Laurin et al. (2019) were able to test each of the main competing hypotheses, defined a priori as a branching topology, given their ossification sequence dataset and a likelihood optimality criterion. Their most consistent result comes from their cranial ossification sequences and supports their "LH", or lepospondyl hypothesis (Laurin et al. 2019: fig. 1d). That is, relative to extinct, "amphibian"-grade taxa, Lissamphibia is monophyletic and nested within lepospondyls. Compared to mammals and birds (including dinosaurs), crown amphibian branches of the Tree of Life are exceptionally old. Each lissamphibian clade likely had diverged during Permian times (Marjanovic & Laurin 2008) and the crown group itself may even date to the Carboniferous (Pyron 2011). In contrast to mammoths and moas, no ancient DNA or collagen sequences are going to be available from >300 million-year-old fossils like the lepospondyl *Hyloplesion* (Olori 2013), although recently published methods for incorporating genomic signal from extant taxa (Beck & Baillie 2018; Asher et al. 2019) into studies of fossils could also be applied to these ancient divergences among amphibian-grade tetrapods. Ossification sequences represent another important, additional source of data with which to test the conclusion of Laurin et al. (2019) that monophyletic Lissamphibians shared a common ancestor with lepospondyls, among other hypotheses. **References** Asher, R. J., Smith, M. R., Rankin, A., & Emry, R. J. (2019). Congruence, fossils and the evolutionary tree of rodents and lagomorphs. Royal Society Open Science, 6(7), 190387. doi: [ 10.1098/rsos.190387 ](https://dx.doi.org/ 10.1098/rsos.190387 ) Beck, R. M. D., & Baillie, C. (2018). Improvements in the fossil record may largely resolve current conflicts between morphological and molecular estimates of mammal phylogeny. Proceedings of the Royal Society B: Biological Sciences, 285(1893), 20181632. doi: [ 10.1098/rspb.2018.1632](https://dx.doi.org/ 10.1098/rspb.2018.1632) Laurin, M., Lapauze, O., & Marjanović, D. (2019). What do ossification sequences tell us about the origin of extant amphibians? BioRxiv, 352609, ver. 4 peer-reviewed by PCI Paleo. doi: [ 10.1101/352609](https://dx.doi.org/ 10.1101/352609) Marjanović, D., & Laurin, M. (2008). Assessing confidence intervals for stratigraphic ranges of higher taxa: the case of Lissamphibia. Acta Palaeontologica Polonica, 53(3), 413–432. doi: [ 10.4202/app.2008.0305](https://dx.doi.org/ 10.4202/app.2008.0305) Olori, J. C. (2013). Ontogenetic sequence reconstruction and sequence polymorphism in extinct taxa: an example using early tetrapods (Tetrapoda: Lepospondyli). Paleobiology, 39(3), 400–428. doi: [ 10.1666/12031](https://dx.doi.org/ 10.1666/12031) Pyron, R. A. (2011). Divergence time estimation using fossils as terminal taxa and the origins of Lissamphibia. Systematic Biology, 60(4), 466–481. doi: [ 10.1093/sysbio/syr047](https://dx.doi.org/ 10.1093/sysbio/syr047) Pyron, R. A., & Wiens, J. J. (2011). A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Molecular Phylogenetics and Evolution, 61(2), 543–583. doi: [ 10.1016/j.ympev.2011.06.012](https://dx.doi.org/ 10.1016/j.ympev.2011.06.012) San Mauro, D. (2010). A multilocus timescale for the origin of extant amphibians. Molecular Phylogenetics and Evolution, 56(2), 554–561. doi: [ 10.1016/j.ympev.2010.04.019](https://dx.doi.org/ 10.1016/j.ympev.2010.04.019) Schoch, R. R. (2003). Early larval ontogeny of the Permo-Carboniferous temnospondyl *Sclerocephalus*. Palaeontology, 46(5), 1055–1072. doi: [ 10.1111/1475-4983.00333](https://dx.doi.org/ 10.1111/1475-4983.00333) Schoch, R. R. (2004). Skeleton formation in the Branchiosauridae: a case study in comparing ontogenetic trajectories. Journal of Vertebrate Paleontology, 24(2), 309–319. doi: [ 10.1671/1950](https://dx.doi.org/ 10.1671/1950) Schoch, R. R., & Witzmann, F. (2009). Osteology and relationships of the temnospondyl genus *Sclerocephalus*. Zoological Journal of the Linnean Society, 157(1), 135–168. doi: [ 10.1111/j.1096-3642.2009.00535.x](https://dx.doi.org/ 10.1111/j.1096-3642.2009.00535.x) Smith, K. K. (2001). Heterochrony revisited: the evolution of developmental sequences. Biological Journal of the Linnean Society, 73(2), 169–186. doi: [ 10.1111/j.1095-8312.2001.tb01355.x](https://dx.doi.org/ 10.1111/j.1095-8312.2001.tb01355.x) Tarver, J. E., dos Reis, M., Mirarab, S., Moran, R. J., Parker, S., O’Reilly, J. E., & Pisani, D. (2016). The interrelationships of placental mammals and the limits of phylogenetic inference. Genome Biology and Evolution, 8(2), 330–344. doi: [ 10.1093/gbe/evv261](https://dx.doi.org/ 10.1093/gbe/evv261) Werneburg, R. (2018). Earliest “nursery ground” of temnospondyl amphibians in the Permian. Semana, 32, 3–42. | What do ossification sequences tell us about the origin of extant amphibians? | Michel Laurin, Océane Lapauze, David Marjanović | <p>The origin of extant amphibians has been studied using several sources of data and methods, including phylogenetic analyses of morphological data, molecular dating, stratigraphic data, and integration of ossification sequence data, but a consen... | | Evo-Devo, Phylogenetics, Systematics, Vertebrate paleontology | Robert Asher | 2018-06-22 08:21:31 | ||
22 Sep 2018
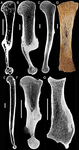
Palaeobiological inferences based on long bone epiphyseal and diaphyseal structure - the forelimb of xenarthrans (Mammalia)Inferences on the lifestyle of fossil xenarthrans based on limb long bone inner structureRecommended by Alexandra Houssaye based on reviews by Andrew Pitsillides and 1 anonymous reviewerBone inner structure bears a strong functional signal and can be used in paleontology to make inferences about the ecology of fossil forms. The increasing use of microtomography enables to analyze both cortical and trabecular features in three dimensions, and thus in long bones to investigate the diaphyseal and epiphyseal structures. Moreover, this can now be done through quantitative, and not only qualitative analyses. Studies focusing on the diaphyseal inner structure (cortical bone and sometimes also spongious bone) of long bones are rather numerous, but essentially based on 2D sections. It is only recently that analyses of the whole diaphyseal structure have been investigated. Studies on the trabecular architecture are much rarer. Amson & Nyakatura (2018) propose a comparative quantitative analysis combining parameters of the epiphyseal trabecular architecture and of the diaphyseal structure, using phylogenetically informed discriminant analyses, and with the aim of inferring the lifestyle of extinct taxa. The group of interest is xenarthrans, one of the four major extant clades of placental mammals. Xenarthrans exhibit different lifestyles, from fully terrestrial to arboreal, and show various degrees of fossoriality. The authors analyzed forelimb long bones of some fossil sloths and made comparisons with several species of extant xenarthrans. The aim was notably to discuss the degree of arboreality and fossoriality of these fossil forms. This study is among the first ones to conjointly analyze both diaphyseal and trabecular parameters to characterize lifestyles, and the first one outside of primates. No fossil form could undoubtedly be assigned to one lifestyle exhibited by extant xenarthrans, though some previous ecological hypotheses could be corroborated. This study also raised some technical challenges, linked to the sample and to the parameters studied, and thus constitutes a great step, from which to go further. References Amson, E., & Nyakatura, J. A. (2018). Palaeobiological inferences based on long bone epiphyseal and diaphyseal structure - the forelimb of xenarthrans (Mammalia). bioRxiv, 318121, ver. 5 peer-reviewed and recommended by PCI Paleo. doi: 10.1101/318121 | Palaeobiological inferences based on long bone epiphyseal and diaphyseal structure - the forelimb of xenarthrans (Mammalia) | Eli Amson & John A. Nyakatura | <p>Trabecular architecture (i.e., the main orientation of the bone trabeculae, their number, mean thickness, spacing, etc.) has been shown experimentally to adapt with great accuracy and sensitivity to the loadings applied to the bone during life.... | | Biomechanics & Functional morphology, Comparative anatomy, Evolutionary biology, Histology, Methods, Morphological evolution, Paleobiology, Vertebrate paleontology | Alexandra Houssaye | 2018-05-14 08:35:20 |




